Созревание (процессинг РНК). Процессинг, сплайсинг. Роль РНК в процессе реализации наследственной информации Процессинг первичных транскриптов у эукариот
Кэпирование и полиаденилирование иРНК называется процессингом (посттранскрип-ционной модификацией).
Кэпирование:
К 5 " концу всех эукариотических иРНК присоединяется во время процессинга остаток 7-метилгуанозина с образованием уникальной 5 "à 5 " фосфодиэфирной связи . Этот дополнительный нуклеотид получил название кэп или колпачек.
Функции кэпа:
1. он защищает РНК от экзонуклеаз
2. помогает связыванию молекулы мРНК с рибосомой.
Полиаденилирование:
3"-конец также модифицируется сразу после завершения транскрипции. Специальный фермент – полиаденилат-полимераза присоединяет к 3"-концу каждого РНК-транскрипта от 20 до 250 остатков адениловой кислоты (поли(А)). Полиаденилатполимераза узнает специфическую последовательность AAУAAA, отщепляет от первичного транскрипта небольшой фрагмент в 11-30 нуклеотидов и затем присоединяет поли(А) последовательность. Принято считать, что такой "хвост" способствует последующему процессингу РНК и экспорту зрелых молекул мРНК из ядра.

По мере участия иРНК в процессах трансляции, длина полиА фрагмента уменьшается. Критическим для стабильности считается 30 адениловых нуклеотидов.
Вся совокупность ядерных транскриптов РНК-полимеразы II известна как гетерогенная ядерная РНК (гяРНК).
Все 3 класса РНК транскрибируются с генов, которые содержат интроны (неинформативные участки)и экзоны (участки ДНК, несущие информацию). Последовательности, кодируемые интронами ДНК, должны быть удалены из первичного транскрипта до того, как РНК станет биологически активной. Процесс удаления копий интронных последовательностей получил название сплайсинга РНК .

Сплайсинг РНК катализируется комплексами белков с РНК , известными как «малые ядерные рибонуклеопротеидные частицы» (мяРНП, англ. small nuclear ribonucleic particles, snRNP ).Такие каталитические РНК носят название рибозимов.
Функции интронов:
· защищают функционально активную часть генома клетки от повреждающего действия химических или физических (лучевых) факторов
· позволяет при помощи так называемого альтернативного сплайсинга увеличить генетическое разнообразие генома без увеличения числа генов.

Альтернативный сплайсинг:
В результате изменения распределение экзонов одного транскрипта во время сплайсинга возникают различные РНК и следовательно различные белки.
Известны уже более 40 генов, транскрипты которых подвергаются альтернативному сплайсингу. Например, транскрипт гена кальцитонина, в результате альтернативного сплайсинга дает РНК, которая служит матрицей для синтеза кальцитонина (в щитовидной железе) или специфического белка, отвечающего за вкусовое восприятие (в мозге). Еще более сложному альтернативному сплайсингу подвергается транскрипт гена -тропомиозина. Были идентифицированы по крайней мере 8 различных тропомиозиновых иРНК, полученных из одного транскрипта (см рис)

33 . Общая схема биосинтеза белка - необходимые предпосылки:
Информационный поток - схема передачи информации (центральная догма молекулярной биологии). Репликация и транскрипция ДНК - ферменты, механизм. Обратная транскрипция, роль ревертаз. Процессинг и сплайсинг иРНК. Характеристика генетического кода, кодон, антикодон.
Отличие биосинтеза белка от биосинтеза других молекул:
· Нет соответствия между числом мономеров матрицы и в продукте реакции (4 нуклеотида--20 аминокислот)
· Между мРНК (матрица) и пептидной цепью белка (продукт) нет комплементарности.
Общая схема биосинтеза белка - необходимые предпосылки:
· информационный поток (передача информации от ДНК на РНК и на белок)
· пластический поток (аминокислоты, мРНК, тРНК, ферменты)
· энергетический поток (макроэрги АТФ, ГТФ, УТФ, ЦТФ)
Первичные транскрипты, образующиеся при транскрипции прокариотических генов, кодирующих белки, функционируют в качестве мРНК без последующей модификации или процессинга. Действительно, трансляция мРНК часто начинается даже до завершения синтеза 3"-конца транскрипта. Совсем иная ситуация наблюдается для молекул рРНК и тРНК. В этом случае кластеры рРНК-или тРНК-генов или даже перемежающиеся участки этих генов часто транскрибируются с образованием единой цепи РНК. И хотя транскрипция этих генов всегда начинается на определенных промоторах и заканчивается на определенных терминаторах, для образования зрелых функциональных форм должны произойти специфическое надрезание первичных РНК-транскриптов и модификация. Подобные молекулярные события называют общим термином посттранскрипционные модификации или просто процессинг РНК. Механизмы процессинга рРНК и тРНК и ферменты, с помощью которых он осуществляется, наиболее полно изучены у Е. coli, и для иллюстрации особенностей посттранскрипционного процессинга РНК мы используем эту систему. Аналогичные модификации эукариотических РНК; в этом случае помимо процессинга рРНК и тРНК используются более сложные системы созревания транскриптов с образованием мРНК.
а. Группы генов, кодирующих рРНК и тРНК
В геноме Е. coli идентифицированы и картированы семь дискретных транскрипционных единиц, кодирующих рРНК. Каждая транскрипционная единица - это молекула РНК, которая состоит из ~5000 нуклеотидов и содержит по одной копии кодирующих последовательностей для 5S-, 16S- и 23S-pPHK. Транскрипция в этой области осуществляется в направлении 16S -> 23S -> 5S. Помимо этих трех последовательностей, кодирующих рРНК, транскрипты содержат вставки разной длины и одну или более копий тРНК-генов. Спейсеры могут находиться перед последовательностями для рРНК, между ними и после них, а тРНК-гены обычно лежат в пределах вкрапленных или 3"-концевых спейсерных сегментов. Для образования функционально зрелых молекул РНК должен произойти процессинг таких транскриптов. До процессинга или во время него происходит модификация специфических оснований в спейсерах, а также в рРНК- и тРНК-генах.
б. Разрезание рРНК-тРНК-котранскриптов
Начальное расщепление первичных транскриптов на фрагменты, содержащие либо тРНК, либо 16S-, 23S- или 58-рРНК-последовательности, осуществляет эндонуклеаза РНКаза III. Ее мишенями служат короткие дуплексы РНК, образующиеся при внутримолекулярном спаривании оснований в последовательностях, фланкирующих каждый из рРНК-сегментов. Например, комплементарные участки в спейсерных областях, фланкирующих последовательность 16S-pPHK, образуют стебель шпильки, в петле которой находится последовательность 16S-pPHK. Аналогичные шпильки образуют и последовательности 23S- и 5S-pPHK. РНКаза III вносит разрывы в двухцепочечный стебель, в результате образуется цепь РНК, содержащая последовательность той или иной рРНК, фланкированную короткими спейсерными участками с 5"-фосфатным и 3"-гидроксильным концами. Затем лишние нуклеотиды спейсерных последовательностей удаляются, возможно с помощью той же самой РНК-экзонуклеазы, которая катализирует и последние этапы процессинга тРНК. В принципе для того, чтобы произошло ферментативное расщепление, должны быть транскрибированы только те нуклеотидные последовательности, которые образуют шпильки. Однако процессинг происходит лишь после завершения синтеза всего первичного транскрипта, поскольку, по-видимому, для правильной укладки целого РНК-транскрипта, который и распознается эндонуклеазой III, необходимы рибосомные или какие-либо другие белки. Процессинг тРНК-сегментов, выщепляющихся из мультигенных транскриптов, осуществляется так же, как и процессинг тРНК из транскрипционных единиц одиночных генов.
в. Образование зрелых тРНК из более крупных транскриптов
Несмотря на то, что некоторые кодирующие тРНК гены находятся внутри транскрипционных единиц рРНК и экспрессируются совместно с генами рРНК, основная часть тРНК-генов представлена одиночными генами или объединена в кластеры. Одни кластеры содержат множественные повторы одних и тех же генов, другие - различные и неродственные тРНК-гены. В некоторых случаях каждый кластер транскрибируется как одна большая молекула РНК, которая подвергается процессингу с последовательным выщеплением зрелых тРНК-фрагментов. Для образования зрелой функциональной тРНК, по-видимому, должны произойти специфическая модификация оснований и присоединение одного, двух или всех трех нуклеотидов 3"-ССА-конца.
Независимо от того, содержит ли первичный транскрипт одну или более тРНК-последовательностей или эти последовательности внедрены в спейсерные участки рРНК, 5"-концы всех тРНК образуются при участии одной эндонуклеазы, называемой РНКазой Р. По-видимому, РНКаза Р узнает характерную свернутую структуру тРНК в полинуклеотиде-предшественнике и отщепляет лидерную или спейсерную последовательности, расположенные перед 5"-концом зрелой последовательности тРНК. 3"-концы тРНК образуются с помощью нескольких активностей. До сих пор неидентифицированная эндонуклеаза расщепляет предшественник в том месте шпильки, где находится 3"-конец зрелой тРНК, а затем другая эндонуклеаза, РНКаза D, завершает образование правильного 3"-конца. В некоторых случаях экзонуклеазное расщепление прекращается точно у 3"-ССА-конца зрелой тРНК, а в других случаях под действием экзонуклеазы образуется конец, служащий затравкой, к которому тРНК-нуклеотидилтрансфераза добавляет один или более инвариантных концевых нуклеотидов.
Отличительной особенностью РНКазы Р является то, что сайт расщепления для нее формируется в результате правильной укладки молекулы тРНК. Изменения в нуклеотидной последовательности, не приводящие к нарушению этой укладки, не сказываются и на процессинге 5"-конца. Другим необычным свойством РНКазы Р является то, что она состоит из белка и РНК. Эта РНК имеет специфическую последовательность из 377 нуклеотидов и сама транскрибируется РНК-полимеразой с гена чуть большего размера и затем подвергается процессингу до размера зрелой молекулы. Удивительной особенностью этой РНК оказалось то, что она одна может катализировать такую же эндонуклеазную реакцию, что и целый рибонуклеопротеин; белок же не обладает самостоятельной эндонуклеазной активностью. Таким образом, эндонуклеазная активность может быть присуща самой РНК, а белок, по-видимому, необходим для сохранения структуры РНК в максимально активной конфигурации.
Зрелые тРНК не только имеют характерную конформацию, но и содержат модифицированные нуклеотиды. Многие из таких модификаций оказываются существенными для выполнения некоторых физиологических функций тРНК. Сегодня охарактеризованы лишь немногие из целой армии ферментов, катализирующих огромное количество реакций модификации. Однако ясно, что модификации происходят в основном на стадии РНК-предшественника и в полностью процессированной тРНК. Такие модифицирующие ферменты представляют особый интерес благодаря своей необычной специфичности в отношении определенных последовательностей: например, только отдельные урациловые остатки превращаются в тиоурацил, метилируются до тимина или восстанавливаются до дигидроурацила. Еще более загадочным представляется образование псевдоуридилата при модификации обычной связи между урацилом и рибозой.
Процессинг - это созревание синтезированной на ДНК преРНК и преобразование её в зрелую РНК. Проходит в ядре клетки у эукариот.
Составные части процессинга
- Удаление нуклеотидов. Результат: значительное уменшение длины и массы исходной РНК.
- Присоединение нуклеотидов. Результат: незначительное увеличение длины и массы исходной РНК.
- Модификация (видоизменение) нуклеотидов. Результат: появление в составе РНК редких "экзотических" минорных ("меньших") нуклеотидов.
Удаление нуклеотидов
1. Отщепление отдельных нуклеотидов по одному с концов цепи РНК. Осуществляется ферментами экзонуклеазами . Обычно преРНК начинается на 5"-конце АТФ или ГТФ, а на 3"-конце заканчивается участками ГЦ. Они нужны только для самой транскрипции, но не нужны для работы РНК, поэтому и отщепляются.
2. Отрезание фрагментов РНК, состоящих из нескольких нуклеоидов. Осуществляется ферментами эндонуклеазами . Таким способом с концов преРНК удаляются спейсерные последовательности нуклеотидов.
3. Разрезание преРНК на отдельные индивидуальные молекулы РНК. Осуществляется ферментами эндонуклеазами. Таким способом получаются рибосомальные РНК (рРНК) и гистоновые (мРНК).
4. Сплайсинг . Это вырезание срединных участков (интронных последовательностей) из преРНК и затем её сшивание . Вырезание осуществляется ферментами эндонуклеазами, а сшивание - лигазами . В результате получается мРНК, состоящая только из экзонных последовательностей нуклеотидов. Все пре-мРНК подвергаются сплайсингу, кроме гистоновых.
В результате удаления нуклеотидов в мРНК может, например, вместо 9200 нуклеотидов остаться всего 1200.
В среднем после процессинга в зрелой мРНК остаётся только 13% от длины пре-мРНК, а 87% теряется.
Присоединение нуклеотидов
К пре-мРНК с начального 5"-конца присоединяется с помощью нетипичной пирофосфатной связи модифицированный 7-метилгуаниловый нуклеотид, это компонент "колпачка" ("шапочки") мРНК. Этот колпачок создаётся ещё на начальном этапе синтеза РНК, для того чтобы защитить нарождающуюся РНК от нападок ферментов-экзонуклеаз, отщепляющих концевые нуклеотиды от РНК.
После завершения синтеза пре-мРНК к её конечному участку со стороны 3"-конца ферментом полиаденилатполимеразой последовательно приращиваются адениловые нуклеотиды, так что получается полиадениловый "хвост" из примерно 200-250 А-нуклеотидов. Мишенью для этого процесса служат последовательности ААУААА и ГГУУУГУУГГУУ в конце преРНК. В результате у преРНК отрезается её собственный хвостик и заменяется на полиА-хвост.
Видео: Снабжение преРНК шапочкой и хвостиком
У пре-тРНК хвост на её 3"-конце создаётся последовательным присоединением трёх нуклеотидов: Ц, Ц и А. Они образуют акцепторную ветвь транспортной РНК.
Модификация нуклеотидов
Важно отметить, что модифицированные минорные нуклеотиды появляются в созревающей РНК именнов в результате процессинга, а не встраиваются в РНК в процессе её синтеза на ДНК.
В нуклеотидах колпачка пре-мРНК происходит метилирование рибозы.
В пре-рРНК метилируются рибозные остатки выборочно по всей длине цепи, с частотой примерно 1%, т.е. 1 нуклеотид из 100.
В пре-тРНК модификация происходит наиболее разнообразно. Например, если уридин восстанавливается, то получается дигидроуридин, если изомеризуется, то получается псевдоуридин, если метилируется, то получается метилуридин, Аденозин может дезаминироваться, превращаясь в инозин, а если затем метилируется, то получается метилинозин. Происходят и другие модификации нуклеотидов.
Видео: Подробно о процессинге
Итог процессинга
Исходные преРНК укорачиваются и модифицируются . В ядре клетки появляются зрелые РНК разных видов: рРНК (28S, 18S, 5,8S, 5S), тРНК (по 1-3 вида для каждой из 20 аминокислот), мРНК (тысячи вариантов в зависимости от количества экспрессированных в данной клетке генов). Здесь же в ядре рРНК связываются с рибосомными белками и формируют большие и малые субъединицы рибосом. Они покидают ядро и выходят в цитоплазму. А мРНК связываются с транспортными белками и в таком виде выходят из ядра в цитоплазму.
Введение
Биосинтез белка можно разделить на стадии транскрипции , процессинга и трансляции . Во время транскрипции происходит считывание генетической информации, зашифрованной в молекулах ДНК, и запись этой информации в молекулы мРНК. В ходе ряда последовательных стадий процессинга из мРНК удаляются некоторые фрагменты, ненужные в последующих стадиях, и происходит редактирование нуклеотидных последовательностей. После транспортировки кода из ядра к рибосомам происходит собственно синтез белковых молекул, путем присоединения отдельных аминокислотных остатков к растущей полипептидной цепи.
Процессинг
Между транскрипцией и трансляцией молекула мРНК претерпевает ряд последовательных изменений, которые обеспечивают созревание функционирующей матрицы для синтеза полипептидной цепочки. С появлением процессинга в эукариотической клетке стало возможено комбинирование экзонов гена для получения большего разнообразия белков, кодируемым единой последовательностью нуклеотидов ДНК.
Кэпирование
Химическая структура кэпа
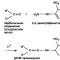
При кэпировании происходит присоединение к 5"-концу транскрипта 7-метилгуанозина посредстом трифосфатного моста, соединяющего их в необычной позиции 5"-5", а также метилирование рибоз двух первых нуклеотидов. Процесс кэпирования начинается еще до окончания транскрипции молекулы пре-мРНК.
Функции кэп-группы:
- регулирование экспорта мРНК из ядра;
- защита 5"-конца транскрипта от экзонуклеаз;
- участие в инициации трансляции
Полиаденилирование
Полиаденилирование заключается в присоединении к 3"-концу транскрипта от 100 до 200 остатков адениловой кислоты, осуществляемом специальным ферментом poly(A)-полимераза.
Сплайсинг
После полиаденилирования мРНК подвергается удалению интронов. Процесс катализируется сплайсосомой и называется сплайсингом.
Трансляция
Готовая белковая молекула затем отщепляется от рибосомы и транспортируется в нужное место клетки . Для достижения своего активного состояния некоторые белки требуют дополнительной посттрансляционной модификации .
Wikimedia Foundation . 2010 .
Смотреть что такое "Процессинг (биология)" в других словарях:
У этого термина существуют и другие значения, см. Процессинг (биология). Процессинг деятельность, включающая в себя обработку и хранение информации, необходимой при осуществлении платежей. Термин часто используется в отрасли банковских… … Википедия
Доставка малых РНК, содержащих шпильки, при помощи вектора на основе лентивируса и механизм РНК интерференции в клетках млекопитающих РНК интерференция (а … Википедия
Пре мРНК со стеблем петлёй. Атомы азота в основаниях выделены голубым, кислорода в фосфатном остове молекулы красным Рибонуклеиновые кислоты (РНК) нуклеиновые кислоты, полимеры нуклеотидов, в состав которых входят остаток… … Википедия
Центральная догма молекулярной биологии обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом… … Википедия
Пре мРНК со стеблем петлёй. Атомы азота в основаниях выделены голубым, кислорода в фосфатном остове молекулы красным Рибонуклеиновые кислоты (РНК) нуклеиновые кислоты, полимеры нуклеотидов, в состав которых входят остаток ортофосфорной кислоты … Википедия
Пре мРНК со стеблем петлёй. Атомы азота в основаниях выделены голубым, кислорода в фосфатном остове молекулы красным Рибонуклеиновая кислота (РНК) одна из трёх основных макромолекул (две другие … Википедия
Центральная догма молекулярной биологии обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом Криком… … Википедия
Схема синтеза белка рибосомой Биосинтез белка сложный многостадийный процесс синтеза полипептидной цепи из … Википедия
Энхансеры.
Усиливающие транскрипцию при взаимодействии со специфическими белками. Энхансеры это не непрерывная – прерывающиеся последовательности ДНК. Они организованы в модули (М1, М2, М3, М4). Одинаковые модули могут встречаться в разных энхансерах, но для каждого энхансера набор модулей уникален. Модуль это короткая последовательность, состоящая не более чем из 2х витков спирали – примерно 20 нуклеотидных пар. Модули ориентированы перед, за и даже внутри гена. Таким образом М1, М2, М3 и М4 это один энхансер состоящий из 4х модулей. Каждый из них узнаётся своими белками, а они в свою очередь взаимодействуют друг с другом. Если в клетке присутствуют все соответствующие белки, то участку ДНК придаётся определённая конформация и начинается синтез мРНК.
Актуализация. Все соматические клетки многоклеточного эукариотического организма имеют одинаковый набор генов. Все гены в них работают на фоновом уровне и не имеют фенотипического проявления, а экспрессируются лишь те, у которых все энхансерные модули узнаны своими белками и эти белки взаимодействуют друг с другом.
Сайленсоры. Это последовательности ослабляющие транскрипцию при взаимодействии с белками. При соответствующем наборе белков экспрессия отдельных генов может быть подавлена.
Некоторые реперссированые (не экспрессирующиеся) гены активируются каскадом событий, запускаемым повышением температуры или синтезом гормона. Гормон, поступив в кровоток, связывается с рецепторами, проникает в клетку, взаимодействует с клеточными белками, изменяет их конформацию, такой белок проникает в ядро, связывается с регуляторным элементом, происходит инициация транскрипции соответствующих генов. Есть белки, которые взаимодействуя с регуляторными элементами блокируют транскрипцию. Например: белок NRSF блокирует транскрипцию соответствующих генов, в нейронах этот белок не синтезируется и как следствие идёт активная транскрипция.
Процессинг РНК у эукариот.
Посттарнскрипционному Ему подвергаются все РНК. Процессинг рРНК и тРНК принципиально не отличается от прокариот.
Процессинг мРНК эукариот
1. Кэпирование. Все 100% синтезированных мРНК. Кэп – метилированый гуанозинтрифосфат присоединенный в необычной позиции (5’ к 5‘)и две метилированые рибозы.
Функции: узнавание кэп-связывающих белков, защита от действии экзонуклеаз
По мере образования про-мРНК (до 30 нуклеотидов) к 5» концу несущему обязательно пурин (аденин, гуанозин) присоединяется гуанин, который затем метилируется. Участие – гуанинтрансферазы.
2. Полиаденилирование. Только 95% всех мРНК и именно эти 95% вступают в этап сплайсинга. Другие 5% не подвергаются сплайсингу и эта матричная РНК в которой зашифрованы альфа и бета интерфероны и белки гистоны.
После завершения синтеза мРНА, полиаденидированию предшествует разрезание специфической эндо кулеазо). Ближе 3» концу про-мРНК, а именно через 20 нуклеотидов после специфической последовательности (ААУАА) синтез безматричный. у каждого вида мРНК полиАхвост определённой длины, покрыт полиАсвязывающими белками. Врея жизни мРНК коррелирует с длиной полиАхвоста.
3. Сплайсингу подвергаются 95% мРНК. Ф. Шарп, 1978 год. Копии вырезанных интронов гидролизуются до нуклеотидов. Осуществляется матюразами. Иногда в сплайсинге участвует sРНК. Правила: 1. фланкированы GT-AG, 2. Нуерация остаётся, но может быть вместе с интронами вырезан экзон.
Цис-сплайсинг (внутримолекулярный сплайсинг) осуществляется в ядре. На первом этапе происходит сборка комплекса сплайсинга. Далее происходит расщепление в 5»сайте сплайсинга, в ходе реакции накапливается два продукта – правильно лигированые экзоны и свободный целый интрон в виде структуры типа «лассо». Множество ядерных факторов белков и рибонуклеопротеидных комплексов - Малые ядерные рибонуклеопротеиды. Этот комплекс, который катализирует сплайсинг, называют сплайсингосомой. Она состоит из интрона, связанного минимум с 5ю мя рнп и некоторыми вспомогательными белками. Сплайсингосомы образуются путём спаривания молекул РНК, присоединением белков к РНК и связыванием этих белков друг с другом. Конечным продуктом такого сплайсинга является вырезание интрона и сшивание фланкирующих его экзонов.
Транс-сплайсинг это пример межмолекулярного сплайсинга. Показан для всех мРНК у трипаносомы и продемонстрирована в опятах ин витро. В ходе него происходит лигирование двух экзонов находящихся в разных молекулах РНК с одновременным удалением фланкирующих их интронов.
Альтернативный сплайсинг обнаружен от дрозофилы до человека и вирусов и показан он для генов, кодирующих белки, участвующие в формировнаии цитоскелета, мышечных сокращений, сборке мемебранных рецепторов, пептидных гормонов, в промежуточном метаболизме и транспозиции ДНК. В сплайсингосоме этот процесс тоже идёт, связан с ферментами занимающимися полиаденилированием. Таким образом мРНК на всём пути следования до завершения трансляции, защищена от нуклеаз с помощью связанных с ней белков (информоферы). Комплекс мРНК с информоферами с ифнормосомы, плюс сРНК. В составе информосом мРНК живёт от нескольких минут до нескольких дней.
4. Редактирование
Сплайсинг тРНК.
Интроны в генах тРНК локализованы через один нуклеотид после антикодона ближе к 3»концу тРНК. От 14 до 60 нуклеотидов. Механизм сплайсинга тРНК лучше всего изучен у дрожжей, а так же в опытах с другими низшими эукариотами и растениями. Задача вырезания интрона в антикодоновой петле реализуется за счёт участия:
Эндонуклеаз (узнать интрон и расщепить про-тРНК в обоих сайтах сплайсинга с образованием свободных 3» и 5»концов экзонов)
Полифункциональный белок (катализирующий все реакции кроме последней – фосфатазной)
2»фосфатаза (удаляет монофосфат с 2»конца 5»концевого экзона)
Лигаза (сшивает)
Сплайсинг рРНК.
Гены ядерных рРНК низших эукариот содержат особые интроны, которые претерпевают уникальный механизм сплайсинга. Это интроны группы I, их нет в генах позвоночных. Общие свойства: сами катализируют свой сплайсинг (автосплайсинг), информация для сплайсинга содержится в коротких внутренних последовательностях внутри интрона(эти последовательности обеспечивают укладку молекулы с образованием характерной пространственной структуры), этот сплайсинг инициируется свободным гуанозином (экзогенным) или любым из его 5»фосфорилированых производных, конечными продуктами являются зрелая рРНК линейная РНК и кор-интроны (кольцевые)
Автоспласинг 1982 г., на инфузория, Томас Чек
Этот процесс чувствителен к ионам магния. Этот сплайсинг показывает что каталитической активностью облажают не только белки но и про-рРНК. Самосплайсинг интронов 1 группы осуществляется последовательно реакций транс-этерификации, где процессы фосфодиэфирного обмена не сопровождаются гидролизом.
Сплайсинг интронов группы 2 мало распространены, обнаружены в 2х митохондриальных генах дрожжей: ген одной из субъединиц цитохромоксидазы и ген цитохрома Б. так же подвергаются самосплайсингу, но инициация сплайсинга и дёт при участии эндогенного гуанозина, то есть гуанозина находящегося в самом интроне. Высвобожденные интроны – подобны лассо, где 5»концевой фосфат РНК интрона соединён фосфодиэфирной связью с 2»гидроксильной группы внутреннего нуклеотида.
Регуляция экспрессии генов у эукариот